

This is going to be old news to readers of this blog who are familiar with the Michael Strauss, Walter Bradley and Guillermo Gonzalez lectures on habitability and fine-tuning. But, it’s nice to see these ideas show up in one of the most prestigious peer-reviewed science journals in the world (if not the most prestigious).
Here’s the article from Science.
It says:
Scientists have known for several years now that stars, galaxies, and almost everything in the universe is moving away from us (and from everything else) at a faster and faster pace. Now, it turns out that the unknown forces behind the rate of this accelerating expansion—a mathematical value called the cosmological constant—may play a previously unexplored role in creating the right conditions for life.
That’s the conclusion of a group of physicists who studied the effects of massive cosmic explosions, called gamma ray bursts, on planets. They found that when it comes to growing life, it’s better to be far away from your neighbors—and the cosmological constant helps thin out the neighborhood.
“In dense environments, you have many explosions, and you’re too close to them,” says cosmologist and theoretical physicist Raul Jimenez of the University of Barcelona in Spain and an author on the new study. “It’s best to be in the outskirts, or in regions that have not been highly populated by small galaxies—and that’s exactly where the Milky Way is.”
Jimenez and his team had previously shown that gamma ray bursts could cause mass extinctions or make planets inhospitable to life by zapping them with radiation and destroying their ozone layer. The bursts channel the radiation into tight beams so powerful that one of them sweeping through a star system could wipe out planets in another galaxy. For their latest work, published this month in Physical Review Letters, they wanted to apply those findings on a broader scale and determine what type of universe would be most likely to support life.
The research is the latest investigation to touch on the so-called anthropic principle: the idea that in some sense the universe is tuned for the emergence of intelligent life. If the forces of nature were much stronger or weaker than physicists observe, proponents note, crucial building blocks of life—such fundamental particles, atoms, or the long-chain molecules needed for the chemistry of life—might not have formed, resulting in a sterile or even completely chaotic universe.
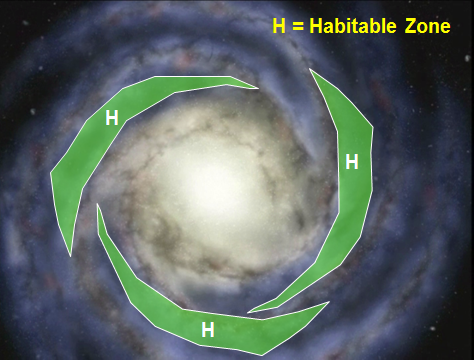
Basically, the best place for a galaxy that permits complex, embodied life to exist is one where you can pick up enough heavy elements from dying stars nearby, but not be in an area that is so crowded by stars that you will be wiped out by intense gamma radiation when they die. So, you want to be between the “arms” of a spiral galaxy, close enough to the areas with a lot of stars, and not too far away. But you can only get that pattern of stars if the universe is expanding at the right rate.
More:
As it turns out, our universe seems to get it just about right. The existing cosmological constant means the rate of expansion is large enough that it minimizes planets’ exposure to gamma ray bursts, but small enough to form lots of hydrogen-burning stars around which life can exist. (A faster expansion rate would make it hard for gas clouds to collapse into stars.)
Jimenez says the expansion of the universe played a bigger role in creating habitable worlds than he expected. “It was surprising to me that you do need the cosmological constant to clear out the region and make it more suburbanlike,” he says.
Remember, this is only one of many characteristics that must obtain in order for a have a location in the universe that can support complex, embodied life of any conceivable kind.
Let’s review the big picture
Time for me to list out some of the things that are required for a galaxy, solar system and planet to support complex embodied life. Not just life as we know it, but life of any conceivable kind given these laws of physics.
- a solar system with a single massive Sun than can serve as a long-lived, stable source of energy
- a terrestrial planet (non-gaseous)
- the planet must be the right distance from the sun in order to preserve liquid water at the surface – if it’s too close, the water is burnt off in a runaway greenhouse effect, if it’s too far, the water is permanently frozen in a runaway glaciation
- the solar system must be placed at the right place in the galaxy – not too near dangerous radiation, but close enough to other stars to be able to absorb heavy elements after neighboring stars die
- a moon of sufficient mass to stabilize the tilt of the planet’s rotation
- plate tectonics
- an oxygen-rich atmosphere
- a sweeper planet to deflect comets, etc.
- planetary neighbors must have non-eccentric orbits
And remember, these requirements for a habitable planet are downstream from the cosmic fine-tuning of constants and quantities that occurs at the Big Bang. No point in talking about the need for plate tectonics if you only have hydrogen in your universe. The habitability requirements are a further problem that comes after the fine-tuning problem.



